
La idea de la localización de funciones en el cerebro se relaciona con el supuesto del enfoque biológico que dice que hay correlatos biológicos en el comportamiento. La teoría de la localización postula que las cogniciones, las emociones y los comportamientos se correlacionan con el funcionamiento de áreas específicas del cerebro. Con el fin de estudiar la localización de las funciones cerebrales los investigadores utilizan técnicas de imágenes cerebrales, autopsias y cirugías; por medio de estas técnicas se investiga la relación entre el comportamiento y los procesos y estructuras cerebrales. Si alguna lesión cerebral afecta una función, se considera que el área lesionada tiene a su cargo esa función específica. Además, la actividad de ciertas áreas cerebrales se ha visto como evidencia de que funciones psicológicas como los comportamientos, pensamientos o emociones están localizadas.
Historia y debate sobre la localización
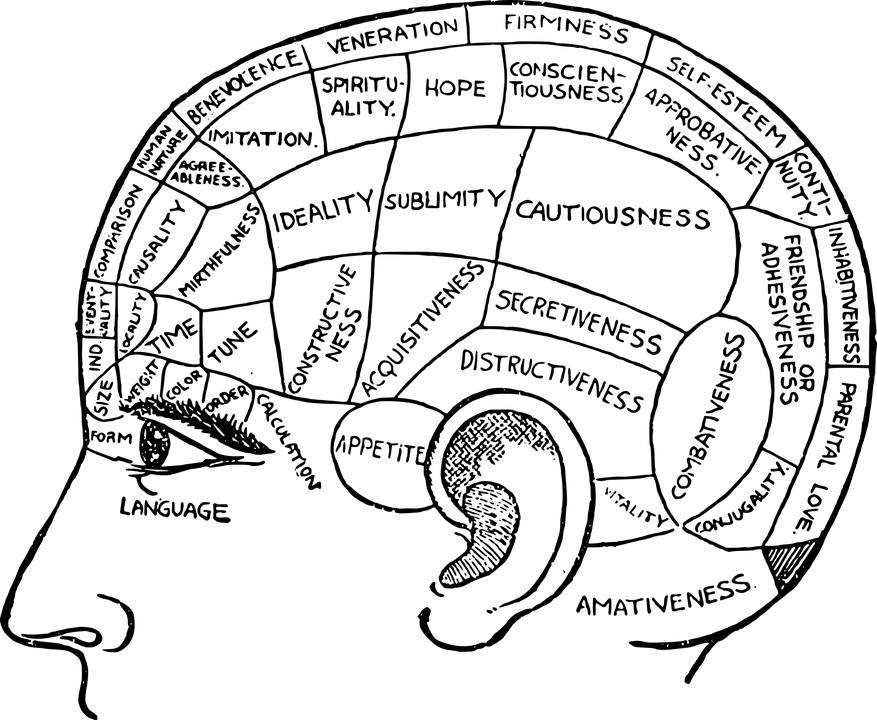
La idea de la localización tiene una larga historia. Este concepto se originó con la frenología del médico alemán Franz Josef Gall (1758-1828). La frenología se considera hoy día como una pseudociencia, la cual proponía que se podía inferir el comportamiento y las capacidades de una persona por la forma del cráneo; proponía que, si un área del cerebro estaba más desarrollada, llevaba a protuberancias en la región correspondiente en el cráneo. Aunque esta idea se ha desacreditado, el supuesto de que ciertas partes del cerebro son responsables de comportamientos específicos todavía es válida. (Luria, 1984)
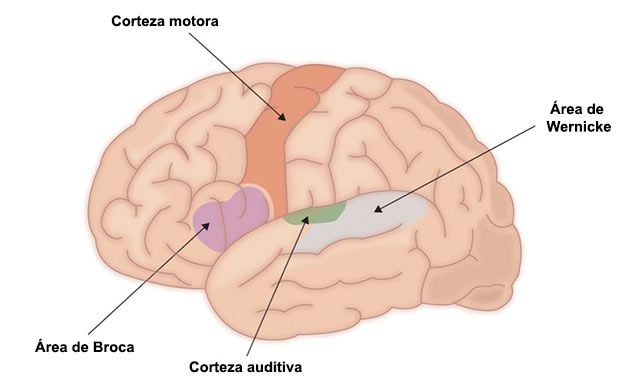
El médico francés Paul Broca (1861) publicó un análisis de caso que culminó en una autopsia, localizó la habilidad para la producción del habla en la mitad posterior de la tercera circunvolución frontal del hemisferio izquierdo, también conocida como giro frontal inferior, del hemisferio izquierdo, ahora conocida como el “área de Broca.” El daño a esta área del cerebro produce una alteración del lenguaje conocida como afasia de Broca. Broca complementó este estudio con 25 casos adicionales (Konnikova, 2013), confirmando sus hallazgos iniciales. Estudios de neuroimagen funcional han mostrado activación del área de Broca en tareas que involucran articulación, confirmando la localización de la articulación del lenguaje en esta área del cerebro (Gazzaniga y colaboradores, 2014).
Esta idea sobre localización de funciones en el cerebro fue complementada con la publicación de Carl Wernicke en 1874 en donde relata lesiones en la mitad posterior de la primera circunvolución temporal, también llamado giro temporal superior, del hemisferio izquierdo del cerebro, hoy día conocida como “área de Wernicke.” Esta lesión lleva a una alteración en la comprensión del lenguaje conocida como afasia sensorial o afasia de Wernicke. Estos estudios de caso muestran que lesiones en áreas diferentes del cerebro perturban de manera diferente el lenguaje, llegando a la conclusión que estas funciones del cerebro están localizadas en áreas específicas del cerebro.
Los estudios de caso de Broca y Wernicke llevaron a un interés en buscar la localización directa de funciones en regiones circunscritas en la corteza cerebral. Como resultado de esto, se diseñaron mapas hipotéticos de la localización de funciones en el cerebro que no tenían el soporte de un análisis psicológico detallado de los síntomas. Estos mapas no diferían mucho de los mapas frenológicos de Gall.
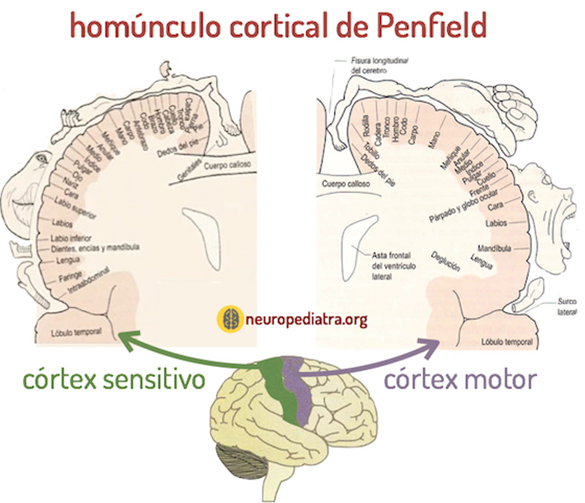
Unos de los mapas de funciones en el cerebro que siguen vigentes son los homúnculos corticales (uno motor y uno sensorial) realizados por Penfield (1981-1976) mediante la estimulación de áreas cerebrales. Mientras realizaba neurocirugías para el tratamiento de la epilepsia, estimulaba diferentes áreas del cerebro y observaba el efecto de esta estimulación en el comportamiento. A partir de estas observaciones, se diseñó un modelo del cuerpo dentro del cerebro que muestra la representación relativa de varias partes del cuerpo en las cortezas motoras y sensoriales. (Gazzaniga y colaboradores, 2014)
En contraposición de esta mirada localizacionista Hughlings Jackson (1835-1911) postuló una hipótesis de que las funciones mentales complejas se deberían analizar desde su nivel de construcción en vez de su localización en áreas específicas del cerebro. Esta hipótesis no fue comprendida hasta mucho tiempo después por otros neurólogos. Para estos especialistas era claro que las funciones elementales tanto sensoriales como motoras se representan en áreas específicas del cerebro, pero dudaban de la aplicabilidad de esta localización estrecha a la actividad mental compleja. Ellos postularon que estas funciones complejas eran el producto de todo el cerebro y no de áreas localizadas de la corteza cerebral. Algunos autores como Karl Lashley (1890-1958) llegaron a proponer que el cerebro era una unidad indiferenciada. (Luria, 1984; Popov y colaboradores, 2017)
La propuesta de Lashley en contra de la localización surgió de sus estudios sobre la memoria con ratas a las cuales les produjo diferentes daños cerebrales. Lashley (1929) hizo experimentos controlados con ratas a las cuales entrenó para navegar en un laberinto con el fin de encontrar comida y luego les produjo una variedad de lesiones en la corteza cerebral, algunas pequeñas y otras más extensas. Luego las volvía a poner en el laberinto y registraba el cambio en el comportamiento. La idea es que, si la memoria estaba localizada, entonces el remover diferentes áreas podría llevar a encontrar la región específica dentro del cerebro responsable de la función de la memoria. Sin embargo, esto no sucedió; lo que observó es que era la cantidad de tejido removido lo que afectaba la adquisición y retención de la memoria en las ratas. Esto lo llevó a concluir que la memoria estaba distribuida en el cerebro en vez de estar localizada. Esto lo llevó a postular dos principios, el de acción de masa y el de equipotencialidad. El principio de acción de masa postula que hay una correlación entre la capacidad para aprender y el porcentaje de cerebro lesionado, no en su localización; el remover más corteza cerebral hacía más lento el aprendizaje independientemente de la localización de la lesión producida. El de equipotencialidad propone que una parte de la corteza puede tomar las funciones de otra zona. Sin embargo, posteriormente se ha observado que la memoria no está distribuida de manera homogénea en la corteza como Lashley pensó. (Luria, 1984; Popov y colaboradores, 2017)
El análisis de esta controversia entre la localización y el holismo, así como de los estudios realizados por Alexander Luria lo llevaron a postular la idea de un sistema funcional complejopara las funciones mentales superiores. Su propuesta postula que los procesos mentales complejos como el pensamiento, el lenguaje, la lectura, la escritura y las matemáticas no se pueden considerar como facultades que estarían localizadas en un área cerebral específica. Por lo tanto, estas actividades conscientes se deben considerar como sistemas funcionales complejos, llevando a una conceptualización diferente de la localización de funciones. Mientras que las funciones elementales pueden tener una localización precisa, las funciones mentales complejas no se podrían localizar en áreas específicas del cerebro sino en sistemas de zonas cerebrales que trabajan de manera coordinada, cada una de las cuales cumple su rol dentro del sistema funcional complejo y pueden estar localizadas en diferentes áreas del cerebro que pueden estar distantes, como en el caso del lenguaje descrito anteriormente y de la memoria que se expondrá más adelante. Esta propuesta de Luria (1984) conlleva también la idea de que la localización de funciones y su organización cerebral puede cambiar a lo largo del desarrollo a medida que el cerebro se va especializando o como resultado de un daño cerebral puesto que la localización no es estática o constante.
Otro aspecto importante relacionado con esta propuesta de Luria (1984) es que una lesión de una de las áreas que conforman el sistema funcional complejo llevaría a la desintegración del sistema y no podría dar razón de su localización. Se tendría que hacer, entonces, un análisis de los síntomas que afectan una función compleja, teniendo en cuenta que lesiones en diferentes áreas del cerebro que se requieren para la ejecución de dicha función la afectarían de manera diferente. Esto se puede ver claramente en el caso de las afasias; la afasia de Broca que afecta la expresión del lenguaje como resultado de una lesión en la mitad posterior de la tercera circunvolución frontal del hemisferio izquierdo es diferente a la afasia de Wernicke que afecta la comprensión del lenguaje debida a una lesión en la mitad posterior de la primera circunvolución temporal. Ambas lesiones llevan a una alteración del lenguaje como función compleja, pero lo hace de manera diferente. Sin embargo, como se describirá más adelante, el lenguaje es más complejo e involucra otras áreas cerebrales y por lo tanto se puede alterar de manera diferente como resultado de diferentes lesiones que forman parte de la red cerebral relacionada con la función de lenguaje. De manera similar se verá posteriormente cómo la memoria también se puede considerar como una función compleja que involucra diferentes zonas cerebrales, cada una de las cuales cumple con un rol diferente en la memoria.
Por su parte, Llinás (2003) señala su preocupación a la mirada que se ha tenido sobre la localización de funciones en el cerebro. Considera que se ha tenido una mirada simplista de la organización modular, similar a la frenología, la cual ha sido sustentada por las técnicas de neuroimagen, llevando a una neofrenología. El cerebro tiene una habilidad limitada para reorganizarse por lo cual la localización de funciones puede ser imprecisa, además de la posibilidad de que las funciones migren de una parte del cerebro a otra como resultado de la neuroplasticidad en casos de lesiones cerebrales. Sin embargo, la dificultad para señalar un evento cerebral de unos pocos centímetros cúbicos no es suficiente evidencia para descartar totalmente la noción de modularidad, en especial si esa modularidad se considera una estructura funcional.
En la actualidad las neurociencias apoyan una idea similar a la de Luria; admite la localización para algunas funciones, pero con limitaciones. Se acepta que funciones elementales tanto sensoriales como motoras se pueden localizar de manera estricta en el cerebro, pero en el caso de las funciones complejas varias áreas cerebrales pueden ser responsables para su ejecución, algunas de las cuales tendrían más importancia que otras. También aceptan que algunas funciones se distribuyen extensamente en el cerebro que se conectan mediante redes neuronales; los estudios que se están llevando a cabo en el Proyecto Conectoma busca tener un mayor conocimiento de estas redes neuronales. Adicionalmente, se ha propuesto que la localización no es estática sino dinámica debido a la neuroplasticidad.
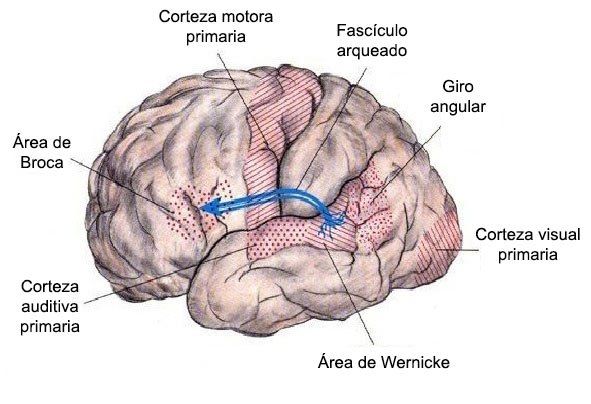
Conectando lo anterior con estudios recientes sobre el lenguaje, se ha llegado a la conclusión que las áreas de Broca y de Wernicke no serían las únicas dos áreas relacionadas con funciones de lenguaje. Esto lo evidencia el análisis hecho por Dronkers y colaboradores (2007) a dos cerebros de los pacientes de Broca cuyos cerebros fueron preservados y a quienes les hizo RMEs. Este análisis mostró que el fascículo arqueado y otras áreas de la corteza cerebral también están relacionadas con el lenguaje. Adicionalmente, Tremblay y Dick (2016) han postulado la idea de que el lenguaje está distribuido en varias áreas cerebrales que están conectadas por diversos fascículos o vías de fibras neuronales, siendo un sistema mucho más complejo e integral que el modelo clásico que comprende las áreas de Broca y Wernicke, conectadas por el fascículo arqueado. Existe mucha evidencia sobre esta propuesta relacionada con la gran complejidad del lenguaje, pues comprende una gran red de áreas cerebrales que lo soporta, algunas de ellas del hemisferio derecho que anteriormente no se consideraban que tuvieran un rol en el lenguaje. De esta manera se ha propuesto que el procesamiento del lenguaje parece elaborarse de manera diferente en los dos hemisferios; en el izquierdo se procesa la expresión y comprensión del lenguaje mientras que en el derecho los aspectos emocionales de éste relacionados con la prosodia y la comprensión emocional del mensaje (Gazzaniga y colaboradores, 2014), lo cual se ha evidenciado en estudios de neuroimagen funcional. Se podría decir, entonces, que el lenguaje, siendo una función compleja, requiere de la actividad coordinada de varias zonas cerebrales y que el daño causado a una de ellas va a alterar el lenguaje de una manera diferente.
En relación con la memoria, el estudio de caso de Henry Molaison, conocido a lo largo de su vida como HM, comenzó a revelar que diferentes aspectos de esta función se relacionan con diferentes áreas cerebrales. HM presentó una severa amnesia anterógrada y una amnesia retrógrada parcial como resultado de una cirugía para controlar la epilepsia, cirugía realizada por el neurocirujano Scoville, quien le retiró los hipocampos y otras áreas de los lóbulos temporales mediales. La amnesia anterógrada afecta la creación de nuevas memorias explícitas, tanto episódicas (memoria para eventos que suceden en un tiempo y lugar específicos) como semánticas (conocimiento general sobre el mundo). La amnesia retrógrada se relaciona con la dificultad para recuperar memorias del pasado, antes de que ocurra un daño cerebral como en el caso de HM.
De este estudio de caso, el cual llevó a cabo a lo largo de más de 50 años, se han hecho varias publicaciones como la de Scoville y Milner (1957), Milner y colaboradores (1968) y Corkin (1968, 1984, 2002 y 2013), entre otras, además del de Corkin y colaboradores (1997), quienes le hicieron una RME. El análisis hecho a HM mostró que el hipocampo y las zonas aledañas del lóbulo temporal medial, afectan la posibilidad de almacenar en la memoria a largo plazo las memorias explícitas o declarativas como la episódica y la semántica, sin alterar la posibilidad de crear nuevas memorias implícitas como la memoria procedimental. La memoria episódica se refiere a aquella relacionada con las experiencias que cada uno tiene a lo largo de su vida y la memoria semántica comprende aprendizajes que uno tiene como resultado de lo que le han contado y lo que ha leído o estudiado. Estas memorias declarativas son la columna vertebral de nuestra vida ya que permiten adquirir los conocimientos que necesitamos para alcanzar logros y sueños, así como para funcionar como personas independientes y dependen de los lóbulos temporales mediales (Corkin, 2013). La memoria procedimental, por otro lado, se relaciona con procesos como montar en bicicleta o conducir que tienen una base motora y se automatizan, la cual se relaciona con los ganglios basales. Por otro lado, la información semántica ya almacenada en la memoria a largo plazo se almacena en áreas de la corteza cerebral, pudiéndose recuperar sin la necesidad de los lóbulos temporales mediales. Sin embargo, Corkin (2013) consideró que las memorias episódicas requieren de la participación continua de los lóbulos temporales mediales porque no solamente participa en su codificación y almacenamiento sino también en su recuperación.
Estudios como los de Maguire y colaboradores (2000, 2006), el de Woollett y Maguire (2011) y el de Antonova y colaboradores (2011) muestran también la importancia del hipocampo en la memoria espacial, especialmente el hipocampo posterior derecho. También evidencian que la memoria involucra varias áreas del cerebro con el fin de procesar diferentes aspectos relacionados con la memoria, siendo ésta una colección de muchos procesos diferentes y no uno sólo. Por lo tanto, nuestros cerebros llevan a cabo diferentes procedimientos relacionados con la memoria de manera simultánea en la medida que forma, consolida y recupera diferentes tipos de memorias, haciendo uso de una variedad de circuitos cerebrales especializados. (Corkin, 2013)
Adicionalmente, estudios en las etapas tempranas en pacientes con Alzheimer muestran que el hipocampo es una de las primeras zonas cerebrales afectadas, llevando a problemas con la memoria reciente y en la orientación espacial. Como se vio en el estudio de Antonova y colaboradores (2011) en esta zona hay receptores de acetilcolina, neurotransmisor cuyo rol en la memoria y el aprendizaje es importante. Daños en el hipocampo que tienen receptores de acetilcolina se han observado en etapas tempranas de la enfermedad de Alzheimer, llevando a los problemas de memoria que se observan en estas etapas de la enfermedad (Jahn, 2013).
Se puede concluir, entonces, que las funciones elementales como algunas de tipo sensorial y motor pueden estar localizadas en áreas específicas del cerebro y el daño en una de estas áreas lleva a una pérdida de la función relacionada. Sin embargo, las funciones más complejas dependen de redes neuronales y por lo tanto de una variedad de áreas cerebrales que pueden estar localizadas de manera cercana o lejana, configurándose en un sistema funcional complejo como lo propuso inicialmente Luria (1984). Teniendo en cuenta esto, lesiones específicas del cerebro afectarían una función compleja de manera diferente, dependiendo de la zona afectada. Igualmente, estudios de neuroimagen evidencian activación de diversas áreas cerebrales al ejecutar tareas complejas. Por último, es importante tener en cuenta que la localización de funciones no es estática debido a neuroplasticidad, de tal manera que las funciones se van especializando a lo largo del desarrollo y en casos de daño cerebral, debido a que algunas áreas cumplen con varias funciones, éstas pueden adquirir las funciones de áreas que se han lesionado.
Recomendaciones para la evaluación:
Aunque es importante que los estudiantes tengan conocimiento de una variedad de ejemplos de localización de funciones en el cerebro, al prepararse para una evaluación se recomienda que enfaticen uno de éstos y por lo menos dos estudios relacionados con el funcionamiento de un área cerebral. Deben poder responder preguntas como las siguientes:
Pregunta para respuesta corta:
- Analice la localización de una función en el cerebro utilizando un estudio relevante.
Preguntas para respuestas largas tipo ensayo:
- Evalúela teoría de localización de funciones en el cerebro, utilizando investigaciones para sustentar su respuesta.
- Discuta uno o más estudios sobre la localización de funciones en el cerebro.
¿En qué medida se puede explicar el comportamiento mediante la localización de funciones en el cerebro?
Limitaciones del concepto de localización de funciones en el cerebro:
- Hay una variabilidad en la anatomía y fisiología del cerebro entre diferentes personas tanto por predisposición genética como por la neuroplasticidad.
- Los estudios sobre la localización han llevado a posturas reduccionistas en las cuales comportamientos específicos se relacionan con áreas específicas del cerebro y por lo tanto asumen que estas áreas son las causantes de esos comportamientos.
- Debido a la variabilidad anatómica y fisiología del cerebro, no se puede concluir de manera inequívoca una relación causa-efecto entre un área cerebral específica y una función psicológica.
- Las funciones complejas requieren de un análisis detallado no sólo de las diferentes áreas cerebrales involucradas sino también de factores socioculturales y cognitivos que influyen en el comportamiento humano.
- El cerebro es un órgano complejo y altamente interconectado y por lo tanto muchas funciones, especialmente las más complejas, requieren de la activación de una variedad de áreas cerebrales que deben trabajar de manera coordinada.
- Los estudios de imagen cerebral que investigan la relación entre la anatomía y funcionamiento del cerebro con el comportamiento todavía carecen de una adecuada resolución para evidenciar la complejidad del funcionamiento cerebral. Por este motivo, en ocasiones diferentes estudios muestran diferencias en la relación entre áreas cerebrales y el comportamiento.
Bibliografía:
Antonova, E., Parslow, D., Brammer, M., Simmons, A., Williams, S., Dawson, G.R. y Morris, R. (2011). Scopolamine disrupts hippocampal activity during allocentric spatial memory in humans: An fMRI study using a virtual reality analogue of the Morris Water Maze. Journal of Psychopharmacology, 25(9), 1256-1265.
Broca, P. (1861). remarks on the seat of the faculty of articulated language, following an observation of aphemia (loss of speech). Bulletin de la Société Anatomique, 6, 330-357.
Corkin, S. (1968) Acquisition of motor skill after bilateral medial temporal-lobe excision. Neuropsychologia, 6, 255-265.
Corkin. S. (1984) Lasting consequences of bilateral medial temporal lobe lobectomy: Clinical course and experimental findings in H.M. Seminars in Neurology, 4(2), 249-259.
Corkin, S. (2002) What’s new with the amnesic patient H.M.? Nature Reviews Neuroscience, 3(2), 153–160.
Corkin. S. (2013) Permanent present tense: The unforgettable life of amnesic patient H.M. New York: Basic Books.
Corkin, S., Amaral, D.G., González, R.G., Johnson, K.A. y Hyman, B.T. (1997). H. M.’s medial temporal lobe lesion: Findings from magnetic resonance imaging. The Journal of Neuroscience, 17(10), 3964-3979).
Dronkers, N.F., Plaisant, O., Iba-Zizen. M.T. y Cabanis, E.A. (2007). Paul Broca’s historic cases: high resolution MR imaging of the brains of Leborgne and Lelong. Brain, 130, 1432-1441.
Gazzaniga, M.S., Ivry, R.B. y Mangun, G.R. (2014. Cognitive Neuroscience: The Biology of the Mind (4a. Edición). New York. W.W. Norton & Company.
Jahn, H. (2013) Memory loss in Alzheimer’s disease. Dialogues in Clinical Neuroscience. 15(4), 445-454.
Konnicova, M. (2013). The man who couldn t speak and how he revolutionized psychology. Scientific American.
Llinás, R.R. (2003) El cerebro y el mito del yo. Bogotá: Editorial Norma.
Luria. A.R. (1984) El cerebro en acción. Barcelona: Martínez Roca
Maguire, E.A., Gadian, D.G., Johnsrude, I.S., Good, C.D., Ashburner, J., Frackowiak, R.S.J. y Frith, C.D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Science, 97(8), 4398-4403.
Maguire, E.A., Woollett, K. y Spiers, H.J. (2006). London taxi drivers and bus drivers: A structural MRI and neuropsychological analysis. Hippocampus, 16, 1091-1101.
Milner, B. y Klein, D. (2006) Loss of recent memory after bilateral hippocampal lesions: memory and memories—looking back and looking forward. Journal of Neurology, Neurosurgery and Psychiatry, 87(3), 230.
Milner, B., Corkin, S. y Teuber, H.L. (1968). Loss of recent memory after bilateral hippocampal lesions syndrome: 14-year follow-up study of H.M. Neuropsychologia, 6, 215-234.
Popov, A., Parker, L. & Seath, D. (2017) IB Psychology course companion (2nd Edition). Oxford: Oxford University Press.
Scoville, W.B. y Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery and Psychiatry, 20, 11-21.
Tremblay, P. y Dick, A.S. (2016). Broca and Wernicke are dead, or moving past the classic model of language neurobiology. Brain & Language, 162, 60-71.
Wernicke, C. (1874/1969). The symptom complex of aphasia: A psychological study on an anatomical basis. En R. S. Cohen y M. W. Wartofsky (Eds.) Boston Studies in the Philosophy of Science, IV, New York: Humanities Press.
Woollett, K. y Maguire, E. (2011). Acquiring “the knowledge” of London’s layout drives structural brain changes. Current Biology, 21(24), 2109–2114.